- عضویت
- 2017/05/18
- ارسالی ها
- 35,488
- امتیاز واکنش
- 104,218
- امتیاز
- 1,376
سلول عصبی
مقدمه
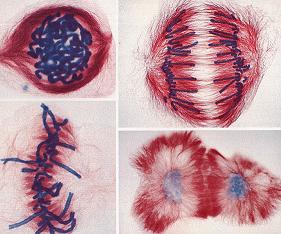
بافت عصبی در واقع از مجموعهای از سلولهای عصبی یا نورون و سلولهای گلیا تشکیل شده است. نورونها دارای اشکال و اندازههای مختلفی میباشند. با این حال هر نورون از سه قسمت پریکاریون ، اکسون و دندریت تشکیل شده است. اکسون غالبا استطاله واحدیست ولی گاهی دارای زواید جانبی به نام کلاترال میباشد. بر اساس نقطه خروج زواید از پریکاریون میتوان نورونها را به یک قطبی ، دو قطبی و چند قطبی تقسیم نمود. نورونهای یک قطبی را میتوان در گانگلیونهای نخاعی مشاهده نمود.

نورونهای دو قطبی اکسون و دندریتها از دو نقطه متقابل از پریکاریون خارج میگردند. این گونه نورونها در لایه میانی قشر مخچه یافت میشوند. پریکاریون آنها بطری شکل است از یک قطب آن دندریت واحدی خارج میشود و سپس به شکل شاخههای درخت منشعب میگردد.
پریکاریون
اندازه پریکاریونها متفاوت است (بین 4 الی 135 میکرون) اشکال آن کروی ، ستارهای ، هرمی و گاه گلابی است. احتمالا در قشر مخ تنها 14 میلیارد نورون وجود دارد. نورونها معمولا دارای هستههای درشتاند که درون آنها یک یا چند هستک وجود دارند. نورونها علاوه بر اندامکهای متداول سیتوپلاسمی مانند دستگاه گلژی ، میتوکندری دارای ویژگیهایی نیز میباشند مانند وجود ذرات نیسل در پریکاریون و دندریتهاست که اجسامی بازوفیلیک محسوب میشوند.
مقدار این ذرات در سلولهای حرکتی بیش از سلولهای حسی است. ویژگی دیگر وجود تارهای نوروفیبریل در سیتوپلاسم دندریتها و اکسونهاست. اعتقاد بر این است که نورفیبریلها به انتقال مواد مثل یونها و متابولیتها کمک میکنند. نورونها گاه محتوی رنگدانههایی میباشند مانند آهن در هسته گلوبوس پالیدوس و همچنین در هسته قرمز.
جایگاه پریکاریون
نورونها و یا فقط پریکاریون آنها اغلب به صورت متراکم داخل گرههای عصبی یا گنگلیونها قرار دارند. این گرهها هم در سیستم عصبی محیطی به نام گانگلیونهای حسی نخاعی- مغزی و با گنگلیونهای حرکتی احشایی و هم درون سیستم عصبی مرکزی به نام هسته وجود دارند.
گانگلیونها
اندازه گانگلیونها و هم چنین تعداد نورونهای هر گانگلیون بسیار متفاوت است (از چند نورون تا 50 هزار).هر گانگلیون درون کپسولی متشکل از بافت همبند محصور میباشد. از کپسول مزبور انشعاباتی به داخل گنگلیونها پیش میروند و آن را به بخشهایی تقسیم میکنند. گانگلیونهای نخاعی را میتوان به صورت برجستگیهای کروی یا دوکی شکل بر سر راه ریشه خلفی اعصاب نخاعی مشاهده نمود. گانگلیونهای خودکار به دو صورت دو ستون فقرات قرار دارند.
تارهای عصبی (اکسونها)
تارهای عصبی در مهرهداران به دو نوع قطور (25 - 1 میکرون) و نازک (کمتر از 1 میکرون) تقسیم میشوند. تارهای قطور به یک یا چند غلاف مجهزاند. معمولا سلولهای شووان تمام اکسونهای محیطی را میپوشانند این سلولها اغلب با یک نظم خاصی همانند شیرینی رولت در اطراف اکسونها پیچیده شدهاند. علاوه بر غلاف شووان ممکن است اکسونها دارای پوششی از چربی تشکیل یافته است منکسر کننده نور میباشد و سفید رنگ است.
گرههای رانویه
میلین در فواصل منظم 0.5 - 0.1 میلیمتر قطع میگردد و بریدگیهایی موسوم به گرههای رانویه را ایجاد میکند. سلولهای شووان که سازنده میلین محسوب میشوند روی میلین قرار میگیرند. تارهای میلیندار فاقد سلولهای شووان در ماده سفید مراکز عصبی فراوان یافت میشوند. از دیدگاه میکروسکوپ الکترونی میلین یک ماده نامنظم و فاقد ساختمان نیست بلکه میتوان آن را به عنوان لایههای متحدالمرکز غشای سلولهای شووان تلقی کرد.
نقش سلولهای شووان
سلولهای شووان در ترمیم اکسونهای قطع شده نقش مهمی را به عهده دارند. هم چنین به هنگام بروز جراحات و آسیب بافتی با عمل فاگوسیتوز (بیگانه خواری) به پاکسازی بافت عصبی میپردازند. عمل سلولهای شووان در داخل سیستم عصبی مرکزی به عهده سلولهای گلیال دیگری به نام الیگودندروسیت میباشد زیرا که دستگاه عصبی مرکزی فاقد سلولهای شووان است. بنابراین عوامل سازنده میلین در سیستم عصبی مرکزی الیگودندروسیتها هستند.
اندازه قطر اکسونها
قطر اکسونها از لحاظ تعیین سرعت انتقال جریان عصبی بسیار مهم میباشند. در سیستم عصبی بایستی پارهای از سیگنالها به سرعت به مراکز منتقل گردند و در برخی حالات نیازی به تسریع انتقال جریان عصبی وجود ندارد. حداکثر سرعت جریان عصبی 120m/sec و حداقل 0.5m/sec میرسد. برای تعیین رابـ ـطه بین سرعت جریان و قطر اکسونها از نوعی تقسیم بندی تارهای عصبی به گروه C,A استفاده میکنند.
گروه A خود دارای چهار زیر گروه α وβ و δ وσ میباشد. این گروه از تارهای میلیندار تشکیل شده است و قطر تارهای آن بین 20 - 1 میکرون نوسان دارد. گروه C فقط شامل یک دسته تار عصبی به قطر 2 - 0.5 میکرون نوسان دارد. گروه C فقط شامل یکدسته تار عصبی به قطر 2- 5/0 میکرون است و در ضمن فاقد میلین میباشد.
خواص نورون
نورونها با داشتن ساختار ویژه برای کاری که انجام میدهند سازگاری حاصل کردهاند. تحـریـ*ک پذیری ، هدایت پیام عصبی و انتقال پیام عصبی سه ویژگی عمده نورونها هستند.
تقسیم بندی نورونها
نورونها را بر حسب اینکه پیام عصبی را در چه جهتی هدایت کنند به دو گروه نورونهای حسی ونورنهای حرکتی تقسیم میکنند. نورونهای حسی دندریت بلند و اکسون کوتاه دارند و پیامهای عصبی را از گیرندههای حسی به سمت مراکز عصبی هدایت میکنند و نورونهای حرکتی اکسون بلند و دندریت کوتاه دارند و پیامها را به اندامهای واکنش مانند ماهیچهها و غدهها میرسانند.
بافت همراه نورون
سیستم عصبی علاوه بر نورون شامل سلولهای دیگری است که بر اعمال حفاظتی و تغذیهای نورون شرکت دارند اما در نقل و انتقال جریان عصبی دخالت مستقیمی ندارند. بافت همراه یا نوروگلی عامل پیوند اجزای متشکل بافت عصبی در سیستم مرکزی و محیطی است. نوروگلی را میتوان به سلولهای درشت و یا ریز تقسیم نمود. از جمله سلولهای درشت آستروسیتها و الیگودندروسیتها هستند. آستروسیتها در ترمیم و استحکام بافت عصبی و جدا نمودن تارها از هم و متابولیسم نقش دارند و الیگودندروسیتها در تولید میلین نقش دارند و سلولهای ریز نوروگلی خاصیت بیگانه خواری دارند.
مقدمه
بافت عصبی در واقع از مجموعهای از سلولهای عصبی یا نورون و سلولهای گلیا تشکیل شده است. نورونها دارای اشکال و اندازههای مختلفی میباشند. با این حال هر نورون از سه قسمت پریکاریون ، اکسون و دندریت تشکیل شده است. اکسون غالبا استطاله واحدیست ولی گاهی دارای زواید جانبی به نام کلاترال میباشد. بر اساس نقطه خروج زواید از پریکاریون میتوان نورونها را به یک قطبی ، دو قطبی و چند قطبی تقسیم نمود. نورونهای یک قطبی را میتوان در گانگلیونهای نخاعی مشاهده نمود.
نورونهای دو قطبی اکسون و دندریتها از دو نقطه متقابل از پریکاریون خارج میگردند. این گونه نورونها در لایه میانی قشر مخچه یافت میشوند. پریکاریون آنها بطری شکل است از یک قطب آن دندریت واحدی خارج میشود و سپس به شکل شاخههای درخت منشعب میگردد.
پریکاریون
اندازه پریکاریونها متفاوت است (بین 4 الی 135 میکرون) اشکال آن کروی ، ستارهای ، هرمی و گاه گلابی است. احتمالا در قشر مخ تنها 14 میلیارد نورون وجود دارد. نورونها معمولا دارای هستههای درشتاند که درون آنها یک یا چند هستک وجود دارند. نورونها علاوه بر اندامکهای متداول سیتوپلاسمی مانند دستگاه گلژی ، میتوکندری دارای ویژگیهایی نیز میباشند مانند وجود ذرات نیسل در پریکاریون و دندریتهاست که اجسامی بازوفیلیک محسوب میشوند.
مقدار این ذرات در سلولهای حرکتی بیش از سلولهای حسی است. ویژگی دیگر وجود تارهای نوروفیبریل در سیتوپلاسم دندریتها و اکسونهاست. اعتقاد بر این است که نورفیبریلها به انتقال مواد مثل یونها و متابولیتها کمک میکنند. نورونها گاه محتوی رنگدانههایی میباشند مانند آهن در هسته گلوبوس پالیدوس و همچنین در هسته قرمز.
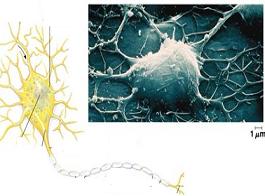
جایگاه پریکاریون
نورونها و یا فقط پریکاریون آنها اغلب به صورت متراکم داخل گرههای عصبی یا گنگلیونها قرار دارند. این گرهها هم در سیستم عصبی محیطی به نام گانگلیونهای حسی نخاعی- مغزی و با گنگلیونهای حرکتی احشایی و هم درون سیستم عصبی مرکزی به نام هسته وجود دارند.
گانگلیونها
اندازه گانگلیونها و هم چنین تعداد نورونهای هر گانگلیون بسیار متفاوت است (از چند نورون تا 50 هزار).هر گانگلیون درون کپسولی متشکل از بافت همبند محصور میباشد. از کپسول مزبور انشعاباتی به داخل گنگلیونها پیش میروند و آن را به بخشهایی تقسیم میکنند. گانگلیونهای نخاعی را میتوان به صورت برجستگیهای کروی یا دوکی شکل بر سر راه ریشه خلفی اعصاب نخاعی مشاهده نمود. گانگلیونهای خودکار به دو صورت دو ستون فقرات قرار دارند.
تارهای عصبی (اکسونها)
تارهای عصبی در مهرهداران به دو نوع قطور (25 - 1 میکرون) و نازک (کمتر از 1 میکرون) تقسیم میشوند. تارهای قطور به یک یا چند غلاف مجهزاند. معمولا سلولهای شووان تمام اکسونهای محیطی را میپوشانند این سلولها اغلب با یک نظم خاصی همانند شیرینی رولت در اطراف اکسونها پیچیده شدهاند. علاوه بر غلاف شووان ممکن است اکسونها دارای پوششی از چربی تشکیل یافته است منکسر کننده نور میباشد و سفید رنگ است.
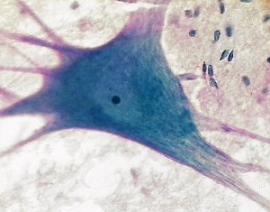
گرههای رانویه
میلین در فواصل منظم 0.5 - 0.1 میلیمتر قطع میگردد و بریدگیهایی موسوم به گرههای رانویه را ایجاد میکند. سلولهای شووان که سازنده میلین محسوب میشوند روی میلین قرار میگیرند. تارهای میلیندار فاقد سلولهای شووان در ماده سفید مراکز عصبی فراوان یافت میشوند. از دیدگاه میکروسکوپ الکترونی میلین یک ماده نامنظم و فاقد ساختمان نیست بلکه میتوان آن را به عنوان لایههای متحدالمرکز غشای سلولهای شووان تلقی کرد.
نقش سلولهای شووان
سلولهای شووان در ترمیم اکسونهای قطع شده نقش مهمی را به عهده دارند. هم چنین به هنگام بروز جراحات و آسیب بافتی با عمل فاگوسیتوز (بیگانه خواری) به پاکسازی بافت عصبی میپردازند. عمل سلولهای شووان در داخل سیستم عصبی مرکزی به عهده سلولهای گلیال دیگری به نام الیگودندروسیت میباشد زیرا که دستگاه عصبی مرکزی فاقد سلولهای شووان است. بنابراین عوامل سازنده میلین در سیستم عصبی مرکزی الیگودندروسیتها هستند.
اندازه قطر اکسونها
قطر اکسونها از لحاظ تعیین سرعت انتقال جریان عصبی بسیار مهم میباشند. در سیستم عصبی بایستی پارهای از سیگنالها به سرعت به مراکز منتقل گردند و در برخی حالات نیازی به تسریع انتقال جریان عصبی وجود ندارد. حداکثر سرعت جریان عصبی 120m/sec و حداقل 0.5m/sec میرسد. برای تعیین رابـ ـطه بین سرعت جریان و قطر اکسونها از نوعی تقسیم بندی تارهای عصبی به گروه C,A استفاده میکنند.
گروه A خود دارای چهار زیر گروه α وβ و δ وσ میباشد. این گروه از تارهای میلیندار تشکیل شده است و قطر تارهای آن بین 20 - 1 میکرون نوسان دارد. گروه C فقط شامل یک دسته تار عصبی به قطر 2 - 0.5 میکرون نوسان دارد. گروه C فقط شامل یکدسته تار عصبی به قطر 2- 5/0 میکرون است و در ضمن فاقد میلین میباشد.
خواص نورون
نورونها با داشتن ساختار ویژه برای کاری که انجام میدهند سازگاری حاصل کردهاند. تحـریـ*ک پذیری ، هدایت پیام عصبی و انتقال پیام عصبی سه ویژگی عمده نورونها هستند.
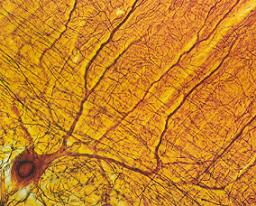
تقسیم بندی نورونها
نورونها را بر حسب اینکه پیام عصبی را در چه جهتی هدایت کنند به دو گروه نورونهای حسی ونورنهای حرکتی تقسیم میکنند. نورونهای حسی دندریت بلند و اکسون کوتاه دارند و پیامهای عصبی را از گیرندههای حسی به سمت مراکز عصبی هدایت میکنند و نورونهای حرکتی اکسون بلند و دندریت کوتاه دارند و پیامها را به اندامهای واکنش مانند ماهیچهها و غدهها میرسانند.
بافت همراه نورون
سیستم عصبی علاوه بر نورون شامل سلولهای دیگری است که بر اعمال حفاظتی و تغذیهای نورون شرکت دارند اما در نقل و انتقال جریان عصبی دخالت مستقیمی ندارند. بافت همراه یا نوروگلی عامل پیوند اجزای متشکل بافت عصبی در سیستم مرکزی و محیطی است. نوروگلی را میتوان به سلولهای درشت و یا ریز تقسیم نمود. از جمله سلولهای درشت آستروسیتها و الیگودندروسیتها هستند. آستروسیتها در ترمیم و استحکام بافت عصبی و جدا نمودن تارها از هم و متابولیسم نقش دارند و الیگودندروسیتها در تولید میلین نقش دارند و سلولهای ریز نوروگلی خاصیت بیگانه خواری دارند.